
A maitriser pour comprendre le cours :
- Définitions de génétique
- Cours de Spécialité de 1ère : Expression de l’IG et transmission de l’IG
I- Le phénotype provient de l’expression du génotype
TP1 – Origine du phénotype + Correction
Les caractères exprimés dans le phénotype sont dus à des protéines, qui sont formées par l’expression des gènes. Différents allèles existent pour un gène, et ils codent pour des protéines différentes donc pour des caractères différents.
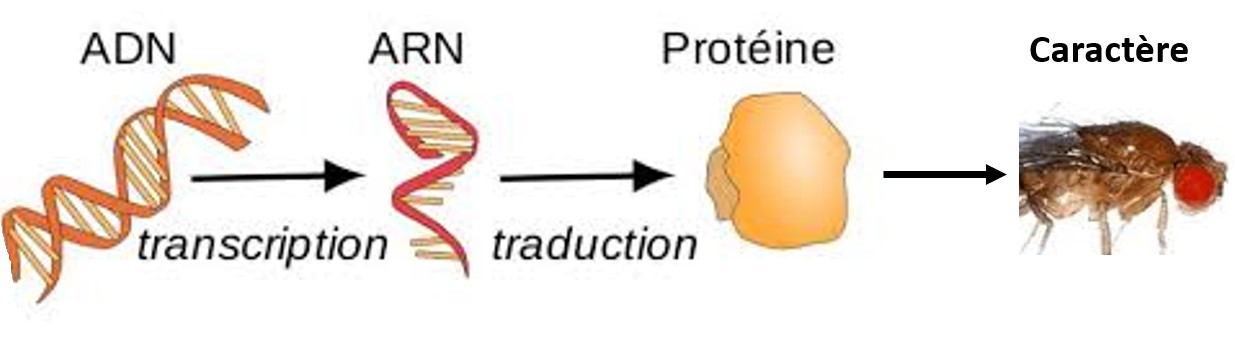
Chez un individu diploïde, chaque cellule contient 2 chromosomes de chaque type, donc 2 exemplaires de chaque gène. Pour un même gène : soit les 2 allèles sont identiques (homozygotie pour ce gène) soit les 2 allèles sont différents (hétérozygotie pour ce gène)
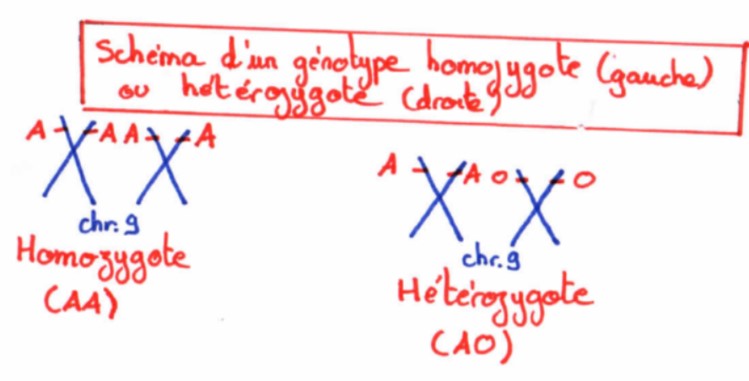
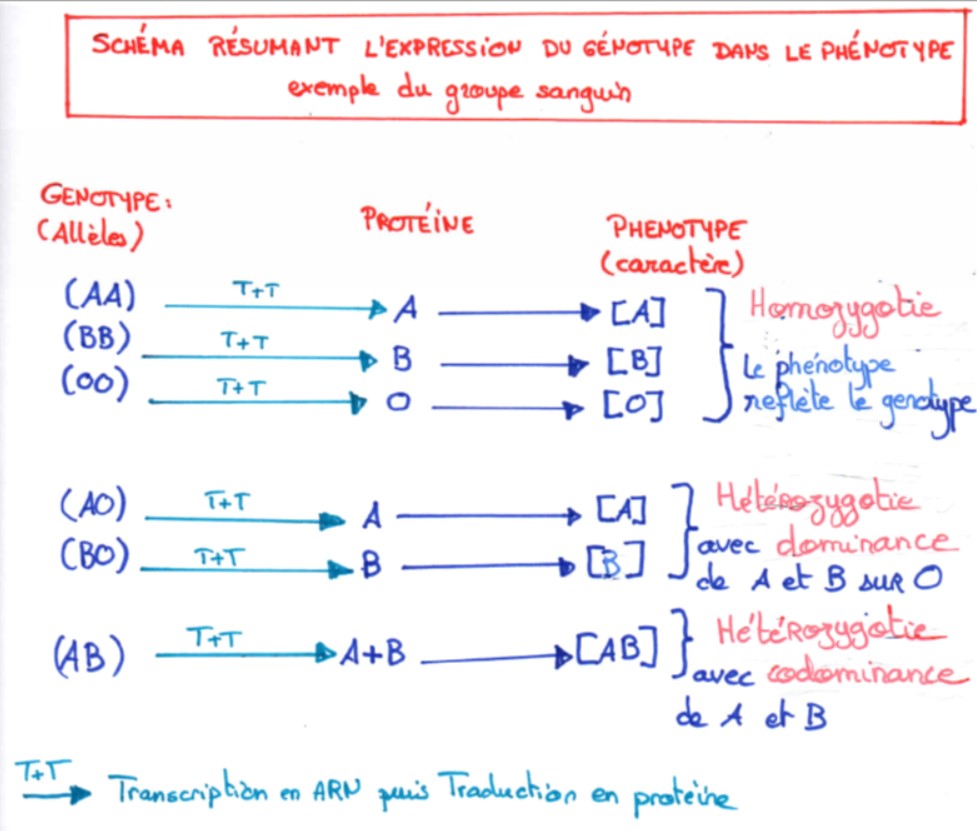
En cas d’homozygotie, le caractère exprimé dans le phénotype est celui de l’allèle présent en double exemplaire
En cas d’hétérozygotie deux cas sont possibles :
- Soit le caractère exprimé dans le phénotype est celui d’un des deux allèles qu’on appellera alors allèle dominant (l’autre est appelé allèle récessif)
- Soit le caractère exprimé dans le phénotype correspond à l’expression des deux allèles, qu’on appellera alors des allèles codominants.
Le phénotype vient de l’expression des allèles présents (donc du génotype) on se demande alors d’où provient le génotype ?
II- Origine du génotype
Lors de la reproduction sexuée, les parents ayant chacun 46 chromosomes fabriquent au sein des organes génitaux (testicules et ovaires) des cellules particulières ayant seulement 23 chromosomes: ce sont les gamètes (ovules/spermatozoïdes/cellules polliniques). Le processus de formation des gamètes est la méiose.
Chaque gamète contient donc un seul allèle de chaque gène. (les gamètes sont donc haploïdes –> toujours)
La fécondation fusionne les génomes des deux gamètes et rétablit la diploïdie : le génotype d’un individu diploïde est donc formé, pour chaque gène, d’un allèle paternel et d’un allèle maternel.
![]()
Cela implique que, si les 2 parents sont homozygotes et ont des allèles différents l’un de l’autre, la descendance aura toujours un allèle venant de chaque parent. Tous les individus de la descendance auront donc le même génotype et le même phénotype : C’est la loi d’uniformité hybride de Mendel.
–> Comment la méiose répartit-elle les allèles dans le cas d’une cellule mère hétérozygote ?
III- Les brassages génétiques lors de la méiose diversifient les génotypes
Lorsque l’on étudie la transmission de plusieurs gènes au cours de la reproduction sexuée, on voit apparaitre de nouveaux génotypes, donc de nouveaux phénotypes. On appelle ces nouveaux génotypes des génotypes recombinés.
Cela implique qu’il y a brassage (= mélange) des allèles des parents.
On va chercher par la suite à étudier les processus qui permettent de produire ces génotypes recombinés (donc de mélanger les allèles parentaux. )
TP 2 – Brassages + Correction
A- Le brassage interchromosomique
A.1- Principe
Le brassage interchromosomique est un mélange d’allèles maternels et paternels dû à la migration aléatoire des chromosomes dans un des gamètes créés.
Lors de la méiose, les chromosomes homologues et donc les allèles qu’ils portent, sont séparés et distribués aléatoirement dans l’un ou l’autre des gamètes créés : cela entraine la formation de nouveaux génotypes.
Cela est dû aux faits :
- Qu’en métaphase 1 : un chromosome à autant de probabilité de se placer au-dessus qu’en dessous du plan équatorial de la cellule : il migre donc aléatoirement dans une des 2 cellules créées
- Qu’en métaphase 2 : un chromatide à autant de probabilité de se placer au-dessus qu’en dessous du plan équatorial : il migre donc aléatoirement dans une des 2 cellules créées
A.2- Conséquences sur la diversité des génotypes
Lorsque l’on étudie deux gènes indépendants, c’est-à-dire situés sur des paires de chromosomes différents, chaque paire de chromosome sépare aléatoirement ses chromosomes dans une des 2 cellules.
Les chromosomes d’origine maternelle peuvent se retrouver entre eux (et donc les paternels seront aussi regroupés), ce qui formera des gamètes ayant des génotypes parentaux (cas n°1 du schéma)
Ou alors, les chromosomes maternels peuvent migrer avec des chromosomes d’origine paternelle, créant des gamètes ayant un génotype recombiné (cas n°2 du schéma)
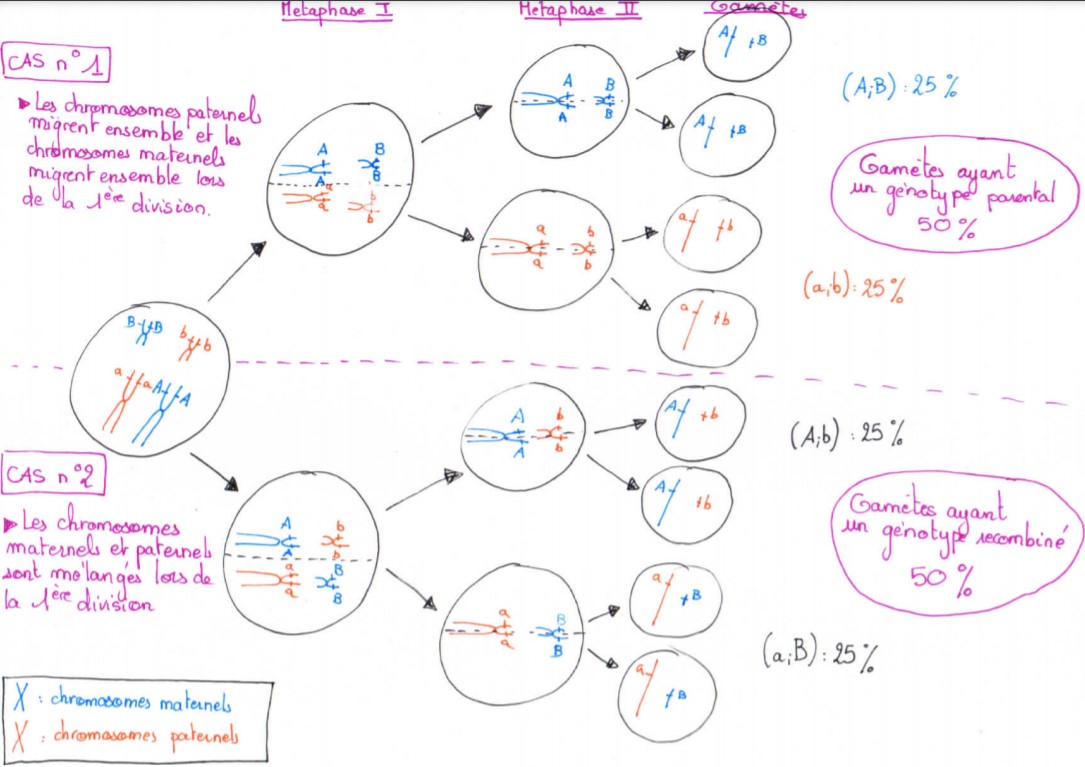
La disposition des chromosomes d’un côté ou de l’autre du plan équatorial étant aléatoire, les proportions de chaque génotype créé seront équivalentes.
Le brassage interchromosomique entraine donc le mélange d’allèles de gènes indépendants, créant ainsi des génotypes recombinés et parentaux dans des proportions équivalentes.
Plus on étudie de gènes indépendants, et plus ils sont à l’état hétérozygote, plus il y a de génotypes possibles par brassage interchromosomique
B- Le brassage intrachromosomique
B.1- Principe
Le brassage intrachromosomique est un mélange d’allèles paternels et maternels situés sur des chromosomes homologues (=de la même paire)
Ce brassage est dû à un phénomène appelé le crossing-over :
Lors de la prophase 1 de méiose, les chromosomes homologues s’apparient et entrecroisent leurs chromatides au niveau de points de croisement appelés les chiasmas.
Dans la majorité des cas, ils se séparent ensuite correctement, mais il arrive (1 à 10% des méioses) que lors de la séparation, les chromatides cassent au niveau du chiasma et que des morceaux de chromatides soient échangés entre les 2 chromosomes homologues. Les allèles portés sur ces chromatides passent alors d’un chromosome à l’autre : c’est le crossing-over.
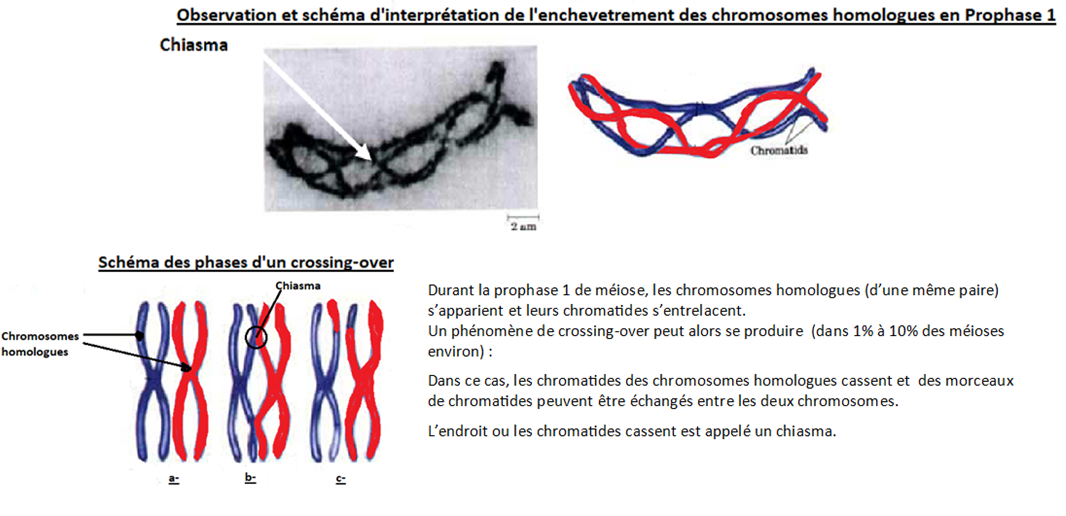
Pour information : C’est le seul cas où vous verrez un chromosome qui portera deux allèles différents sur ses deux chromatides cette année
B.2- Conséquences sur la diversité des génotypes
Si l’on considère alors deux gènes liés, c’est-à-dire portés par le même chromosome : lors d’un crossing-over, il y a mélange des allèles parentaux lors de l’échange des portions de chromatides paternels et maternels, créant des chromosomes recombinés.
Les méioses avec C-O aboutiront donc à la création de gamètes ayant un génotype recombiné
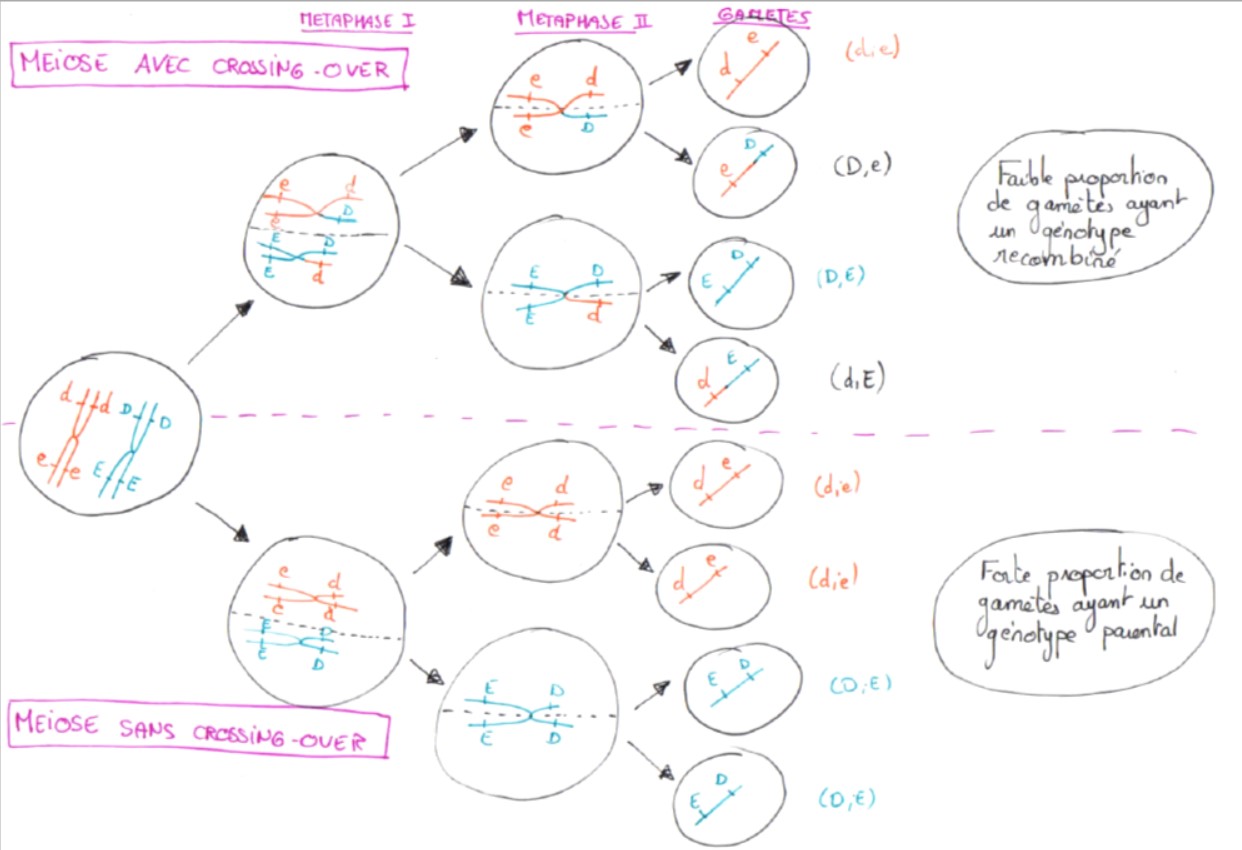
Comme le crossing-over est un évènement rare, la proportion de génotypes recombinés sera plus faible que la proportion de génotypes parentaux.
Le brassage intrachromosomique entraine donc, suite à un crossing-over, le mélange d’allèles de gènes liés, créant ainsi des génotypes recombinés dans de faible proportions.
Les brassages intra et inter créent une diversité quasiment infinie de génotypes de gamètes et la fécondation rassemble ces gamètes de manière aléatoire : les cellules-œufs obtenues auront donc un génotype qui leur est propre
IV- Origine du phénotype des clones et diversification des génotypes sans reproduction sexuée.
TP4 – Origine du phénotype des clones + Correction
Un individu est formé de cellules issues de mitoses successives d’une cellule-œuf initiale. Toutes les cellules issues d’une même mitose constituent un clone et peuvent être isolées (ex : cellules sanguines) ou associées dans un tissu. Elles ont théoriquement toutes le même génome car la mitose est une division conforme.
Cependant, la mitose est précédée d’une réplication, qui copie l’ADN et qui a un taux d’erreur d’environ 1 nucléotide mal copié par milliards de copie : ces erreurs provoquent des mutations qui sont transmises à toutes les cellules issues de la mitose (donc à tout le sous-clone)
Chaque individu est donc constitué d’une mosaïque de sous-clones présentant des différences génétiques liées à ces mutations accumulées au cours de toute la vie de la lignée cellulaire : cela génère de la diversité génétique au sein des cellules d’un même individu.
Ce sont aussi ces accumulations de mutations qui expliquent la diversité génétique observée chez les clones de plantes qui se reproduisent de manière asexuée.
Et la belle vidéo de révision :